







Gouldian Finch head feathers coloring
Gouldian Finch head coloring
Head coloring types
In the wild, three variations of Gouldian Finches exist with natural body coloring. These include red‑headed, black‑headed and yellow (orange)-headed specimens.
It is however not very scientific to say that only three basic species exist. The question is which of these is the most probable nominotypical form, or do we really encounter three different species or subspecies.
The red-headed specimens are considered the nominotypical form living in the wild. The remaining two are true mutations that developed naturally. While it is not impossible for a mutant specimen to survive in the wild, most of the time these birds are clearly visible and helpless and become easy prey for their natural predators. Therefore the mutant species are in danger of becoming extinct. The fact that these mutated Gouldian Finches may be encountered in the wild, may be caused by only a “minor” mutation of head coloring. Birds with only a “minor difference” are barely recognizable among the general population. Breeding examples will show this is not a case of an interspecific cross.

Behavioral and genetic issues
Recent research by Pryke and Griffith described the distinctions in the behavior of red and black-headed Gouldian Finches [Pryke and Griffith, 2009].
In the wild, the black-headed birds are more frequent than the red-headed ones at a 3:1 ratio. It is assumed that further in the past red-headed and black‑headed Finches existed as two geographically separate populations and therefore as two different “subspecies” – in the sense of a single species territorially living in separate communities. Later on they would come into contact again and begin their co-existence as a single population. This original geographic separation may explain why there are such differences present in the behavior of different colored Gouldian Finches.
Both male and female red-headed Finches are more aggressive (vigorous) as the black‑headed ones. The competitive nature of the red‑headed Finches might be explained by more difficult access to nesting cavities, water and food sources in the area, where they lived during the geographic separation.
The “passive” – less competitive nature of the black-headed Finches may be attributed to an environment with plenty of resources, so the birds did not have to fight for food and their character traits contain less genetically coded competitiveness.
Previous research pointed out higher mortality rate of females compared to males in cases when different colored specimens would mate. Pryke and Griffith discovered that when red and black-headed birds mate, less of their offspring survived compared to the offspring of same colored pairs. This finding was however not witnessed in captivity on birds that came from the wild. Even with birds coming from the wild, black-headed birds often chose red-headed partners, even though they could have chosen a partner with the same head color as their own. Breeding between the black and red-headed might be one of the factors contributing to the decline of the wild living population. Under normal circumstances Gouldian Finches prefer mating with birds of the same head color. Black‑headed pairs are able, with certain genetic dispositions to have red‑headed offspring and red-headed pairs are able to have offspring with any of the three head colors. Even though the red-headed birds have a great predisposition advantage of genetic dominance over the black-headed ones (e.g. they occupy the best nesting cavities, get to drink first from the watering-places) they are the minority of the population because of their social aggressiveness, higher susceptibility to stress, higher gonadal hormone levels, etc. These traits are a predisposition to health problems, such as air sac mite infection and due to the higher stress susceptibility they are unable to retain normal immunity level. In the times when numbers of wild-living populations declined dramatically due to capture, most of the females, lacking options for choice an appropriate mating partner, probably mated with different colored males (i.e., red and black-headed mating). The higher count on these different pairs might have had a negative effect on the capability of the local population to grow, due to the reduced probability of youth survival and higher rate of males among the surviving population [Pryke and Griffith, 2009]. The final results showed higher mortality rate of females from such mating [Pryke and Griffith, 2009].
Note 1: Here I would like to point out the fact, that the theory of genetic diversity among red and black-headed birds was not confirmed in captivity. At least not to such an extent that it would be applicable in breeding. I think of breeder experience. I myself also have no such experience. The studies of the population living in the wild were focused on birds that had lived geographically separated for hundreds of generations. By coincidence one of the populations was red‑headed, the other black‑headed. In my opinion, the color of the head itself does not play a role, the important part is, that the red-headed birds lived in a barren environment, which forced them towards competitiveness and therefore towards higher levels of stress. The black‑headed population lived in a more hospitable environment and competitiveness and stress susceptibility had not developed to such extent. If their separation had been long enough, these traits would have become a part of their genetic predispositions and here we might find the roots of red and black‑headed Finch “incompatibility”.
It is a highly probable assumption that the population in the wild still retains these genetically-coded differences. In breeds however, the birds were mated for a long time without concern for head coloring and therefore it may be assumed that the genetic diversity was eliminated, at least to a certain level. In my opinion the genetic set carrying the trait described in the wild population has already mixed sufficiently during domestication. Space remains open for future studies in the wild, as well as in breeds and the question is, of course, open for discussion.
Note 2: In recent times some of the biologists specializing in evolutionary biology expressed a thought that a subspecies, the black-headed Finches, has separated itself from the nominotypical red-headed Gouldian Finch. In their opinion, the current state is a sort of a transition before the development of two independent species. For this it is necessary that the populations are strong enough and that they are separated geographically, though some theories state that a new species may develop in the same territory as the original species, if it specializes. An argument supporting the subspecies theory is the higher mortality of the youth coming from red and black-headed Gouldian Finch pairs (coming from interspecific breeding). Theories have emerged, that a red-headed female picks a red-headed male and thus has more numerous lays, more fertilized eggs and depending on the head-color of the partner the female is able to control the sex of the offspring. In any case I think that these theories are new, unexplored and time will show whether they are correct. Molecular biology might also aid in the ruling, though no ruling is always absolute.
Mating in breeds
Even though I acknowledge foreign studies concerned with the population in the wild, in my breed I have not witnessed any problems when making pairs with different head colors. The mortality rate was no higher or lower whether the parents had same or different colored heads. I see no reason why “pure” head color breeds should be selected. I have noted no problems even when pairs with the same head colors were introduced into the breed during several generations, whether they were red, black or yellow-headed. I have also not experienced that breeding yellow-headed finches across several generations would lead to plumage degradation (flaws). It is however important that the breeder is strictly selective about his breed, so he only introduces birds that are in good physical condition and phenotypically strong. Considering mating birds with different head colors, it is more important what sort of offspring the breeder desires and the pair for mating should be chosen according to this.
Head plumage types
Here we deal with two types of feathers. The plumage of red and yellow‑headed birds is different from the black‑headed ones.
Yellow and red-headed: there are also differences in the structure of the head feathers of males and females
· Male feathers feature long barbs, without barbules with barbicels on the end. Top‑to‑bottom: red part, followed by a brown part and a minute black part. The bottom part of the feather is grayish‑white.
· Female feathers have barbs, without barbules with barbicels on the end, which are quite shorter. Top-to-bottom: red part, followed by a brown part, variable, relatively saturated black part and the bottom is the same as the males’. The female heads typically have a darker tint of red and often yellow due to the structure and the different proportions of the colors of the feather.
These feathers are ornamental in nature and contain red canthaxanthin and yellow lutein (natural carotenoid pigment). If the production of canthaxanthin is interrupted on the head it results in the production of yellow lutein.

Black-headed birds: the head plumage of black-headed males and females is almost identical. The entire vane is covered with barbicels. The feathers are entirely filled with black eumelanin. In other words, compared to the feathers of the red-headed birds these feathers are less developed and lack the “tip”, where the yellow or red pigment is usually located.

Pigments
The plumage of Gouldian Finches contains 4 basic pigments.
Melanin:
· Black (eumelanin). Mostly present in black colored heads.
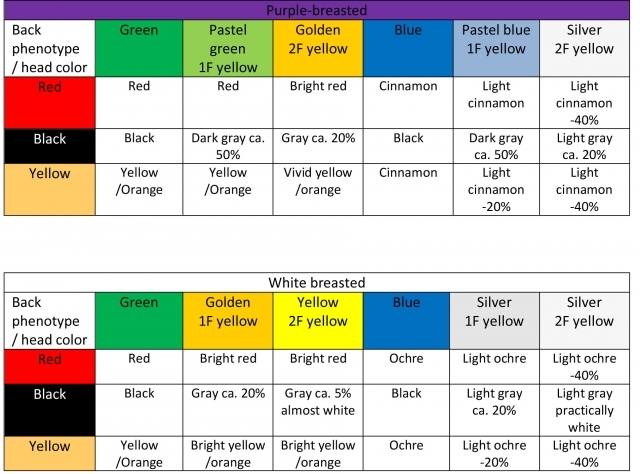
· Brown-red (pheomelanin). Mostly present in the feathers on the breast, where it causes bluish-purple coloring. It facilitates production of structural pigmentation, similar to eumelanin. It is also present in other plumage on the body. It is illustrated on the photos of the heads of yellow-headed birds, where it is obvious that the green purple-breasted and green white-breasted birds have different head coloring. It means that the lack of this pigment determines the absolute intensity of the colors. White-breasted birds have brighter colors.
Carotenoids:
- Yellow (lutein epoxide – alpha xanthophyll). The photo of a yellow‑headed bird illustrates not only the distribution of lutein but also of pheomelanin present in the medulla of the barb.
- Red (canthaxanthin – beta carotenoid). Its intensity may be seen on the photos of red-headed birds. Similarly as in yellow-headed birds, pheomelanin is also present, which results in white-breasted Finches having brighter head coloring.


As may be seen on the displayed picture, the intensity of color in the middle section of the feather “1” is influenced by pheomelanin. In purple-breasted finches it is present, in white-breasted ones it is practically absent. The presence of brown coloring is not dependent on green or double factor yellow coloring of the bird. The section labeled as “2” depends on the mutation of green coloring. In green Finches black eumelanin is present, while it is practically absent in double factor yellows, breast coloring does not affect this part of plumage.

Many color mutations of Gouldian Finches have been bred. Mutation of body color influences the phenotype of head color. Inheritance of the mutations is the same as in naturally colored specimens. Therefore we may speak of red-headed green, blue or silver, even though phenotypically the coloring is different. This is analogically true for other head colorings. In the presented tables I describe the approximate shades present with each mutation. On the photos some color mutations and head coloring differences may be seen. It is mainly important to notice the overall structure of the feathers, the areas with pheomelanin (more prominent in purple-breasted birds) and the area of the lower lateral barbs containing eumelanin depending on the mutation. In green and blue specimens the pigmentation remains the same, in yellows it is lessened.
Abbreviations and symbols
I will abandon my traditional labeling, the one used in the genetic calculator and I will attempt to explain the inheritance on a different, slightly diverse principle. We will assume that red color is dominant and sex linked, black color is recessive and sex linked. In this case we consider black to be a recessive allele to a red dominant allele. The mechanism of pigment production is described elsewhere, so I will mention it only briefly. If, in the process of red pigment – canthaxanthin production, a change occurs, yellow lutein is created. The gene that causes interruption of red pigment production is present in an autosomal chromosome, it is therefore not sex linked and is inherited freely. This gene is a modifier, that is either active and thus disables red pigment production, or is inactive and red pigment is produced.
RR – red head color
rr – black head color
II – active modifier for disabling red pigment production (yellow is produced)
ii – inactive modifier for disabling red pigment production (red is produced)
P – parental – ancestral generation
F1 – 1st generation of offspring
F2 – 2nd generation of offspring
G – gametes (gonadal cells)
Description and properties of the different colors
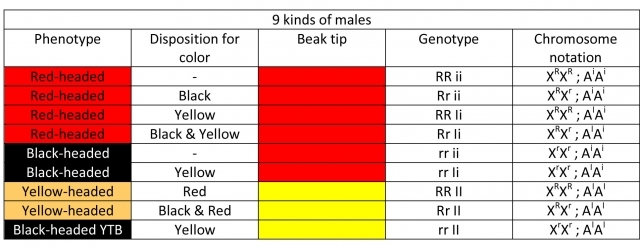
♂ Homozygous males

- Red-headed RR (Red) – have a red-tipped beak: the color is sex linked and dominant.
- Black-headed rr (Black) – have a red-tipped beak: the color is sex linked and recessive. Black head coloring means a lack of red pigment and it seems to be a recessive allele to red. As the color is sex linked and recessive and the males have two sex-related XX chromosomes, they may split to this color.
- Yellow-headed II (Orange) – have a yellow-tipped beak: the genotype manifests only in a double factor state, meaning that both alleles of the modifier have to be active to disable red pigment production.
The modifier for yellow pigment creation is located on an autosomal chromosome, it is inherited unrelated to the sex of the offspring. The yellow has to be present in double factor state to be visible in the phenotype, it is recessive, with the exception of recessive epistasis. For the yellow color to appear in the phenotype of the head, the XX sex‑related chromosomes must contain genes for red head coloring. I call the yellow color recessive, but not in the true meaning of the term. When it is present in a heterozygous – splitting state, it behaves like two different alleles of the same pair relevant to the observed gene – it does not affect the phenotype.
Ii: the genotype contains a single factor of mutated yellow (splits to yellow). Chromosome notation: AIAi
ii: the modifier for yellow pigment creation is inactive. Chromosome notation: AiAi
Black-headed birds with yellow-tipped beak rr;II (YTB). They are genotypically double factor yellow-headed birds with a black head coloring phenotype that prevents expression of the yellow color. It is caused by suppression of yellow lutein by black eumelanin. The yellow modifier is active, but the sex-related chromosomes contain alleles for black coloring.
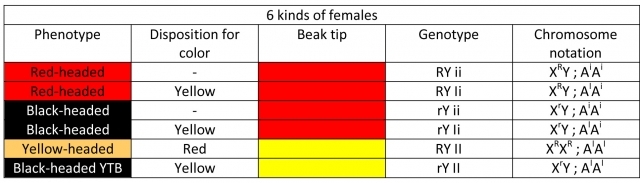
♀ Hemizygous and homozygous females

1. Red-headed RY (Red) – have a red-tipped beak: the color is sex-linked and dominant. Since female are hemizygous for this color, one allele is sufficient for trait manifestation.
2. Black-headed rY (Black) – have a red-tipped beak: the color is sex-linked and recessive to red. Black head coloring means a lack of red pigment and it seems to be a recessive allele to red. As this color is sex-linked and recessive and females have only one sex-related X chromosome, they may not split to this color.
3. Yellow-headed II (Orange) – have a yellow-tipped beak: the genotype manifests only in a double factor state, meaning that both alleles of the modifier have to be active to disable red pigment production.
The modifier for yellow pigment creation is located on an autosomal chromosome, it is inherited unrelated to the sex of the offspring. The yellow has to be present in a double factor state to be visible in the phenotype, it is recessive, with the exception of recessive epistasis. For the yellow color to appear in the phenotype of the head, the XX sex‑related chromosomes must contain genes for red head coloring. I call the yellow color recessive, but not in the true meaning of the term. When it is present in a heterozygous – splitting state, it behaves like two different alleles of the same pair relevant to the observed gene – it does not affect the phenotype.
Ii: the genotype contains a single factor of mutated yellow (splits to yellow). Chromosome notation: AIAi
ii: the modifier for yellow pigment creation is inactive. Chromosome notation: AiAi
- Black-headed birds with yellow-tipped beak rr;II (YTB). They are genotypically double factor yellow-headed birds with a black head coloring phenotype that prevents expression of the yellow color. It is caused by suppression of yellow lutein by black eumelanin. The yellow modifier is active, but the sex-related chromosome contains alleles for black coloring.
These definitions give us 9 types of males and 6 types of females (homo and heterozygous specimens).
Breeding examples
Were these forms diverse species, then we would end up with offspring of hybrids from interspecific mating. Here this is not the case, because if we breed a black-headed male with a red-headed female, we end up with black‑headed females and red-headed males which split to black head coloring.
1st breeding
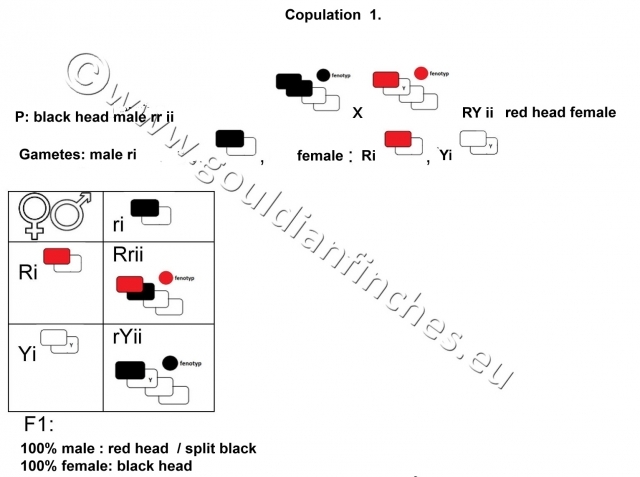
If we breed a red-headed male with a black-headed female, then all offspring will be red-headed, but the males will split to black head coloring. It is very important to know this, because it points to the fact, that the gene responsible for black head coloring and the gene for read plumage color are located on a sex-related chromosome and is therefore sex-linked. The gene for yellow head color is inherited as an autosomal trait, it is located on an autosomal chromosome, it is therefore not sex-linked.
2nd breeding
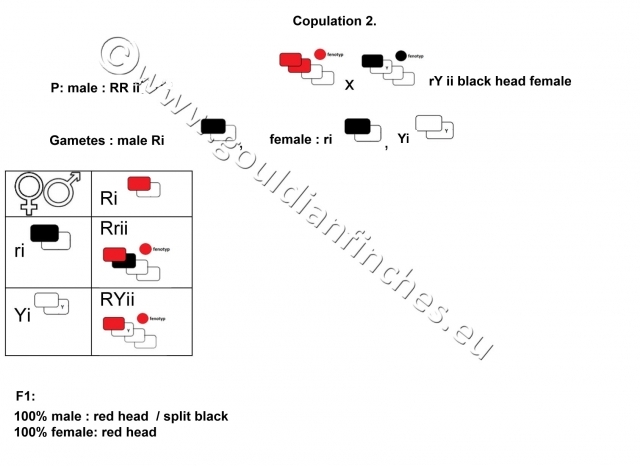
If we breed a yellow-headed male with a black headed female, all of the offspring will be red-headed. All males may split to black and orange head coloring and the females may split to yellow head coloring.
3rd breeding

4th breeding
We breed a black-headed male, splitting to yellow and a yellow-headed female. Both parents carry in their phenotype a recessive coloring, i.e. black and yellow. The offspring will have in their phenotype also the dominant red color.
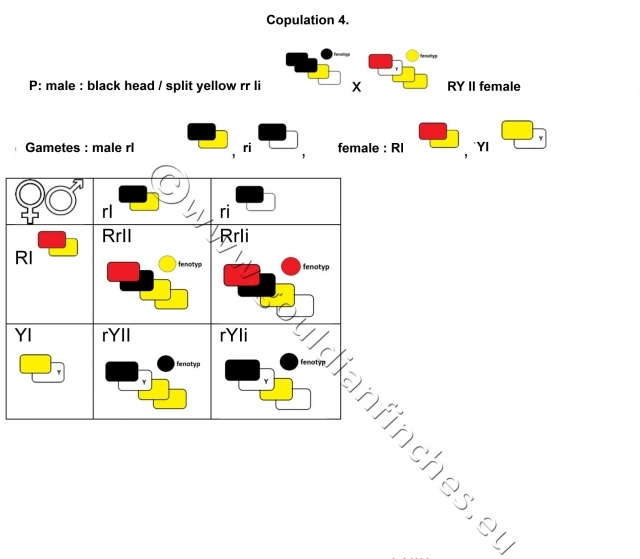
Phenotype F1:
Males:
50% yellow-headed, with genes for both black and red color
50% red-headed, split to black and yellow
Females:**
50% black-headed YTB, with double factor yellow
50% black-headed, split to yellow
The breeding of a black-headed male with a yellow-headed female points to a genetic interaction. The result will be an offspring that has the colors of their parents as well as the dominant red color, which their parents’ phenotypes did not exhibit.
Seemingly, a paradox occurs in this kind of breeding, as if the laws of genetics were broken. From two parents bearing recessive traits in their phenotypes we get an offspring with dominant head coloring, i.e. red-headed. Genetic laws tell us that in principle only the dominant trait – color will manifest in the phenotype and that animals, birds, may not be carriers for the dominant trait unless it manifests in the phenotype. The example of genetic interaction – recessive epistasis – speaks of such a case, when the phenotype manifests a recessive trait, but the dominant trait (via splitting) is still coded in the genotype. Such an example is the head color inheritance in Gouldian Finches.
Recessive epistasis definition: if two recessive alleles are located in a homozygous constitution, they may suppress the phenotypical manifestation of a dominant allele with a different locus in both homozygous and heterozygous constitutions.
This means that the phenotype – the appearance of the offspring may contain also the dominant color that was not observable in the parents’ phenotypes. In other words, birds with a recessive color phenotype may carry a disposition (may split) to a dominant color.
This is the case of the 4th breeding. In the males the double factor of yellow suppressed the red coloring in the phenotype, similarly in females, though they are hemizygous for the red color, they only have one allele for the trait.
Proof, using Mendelian ratios, pointing to recessive epistasis
Characteristic phenotypical ratio of recessive epistasis: 9:3:4,
Phenotypical ratio of recessive epistasis in experimental breed: 1:1:2.
P – parental (ancestral generation)
G – gametes (gonadal cells)
F1 – first filial generation
F2 – second filial generation, parented by F1
Breeding F1
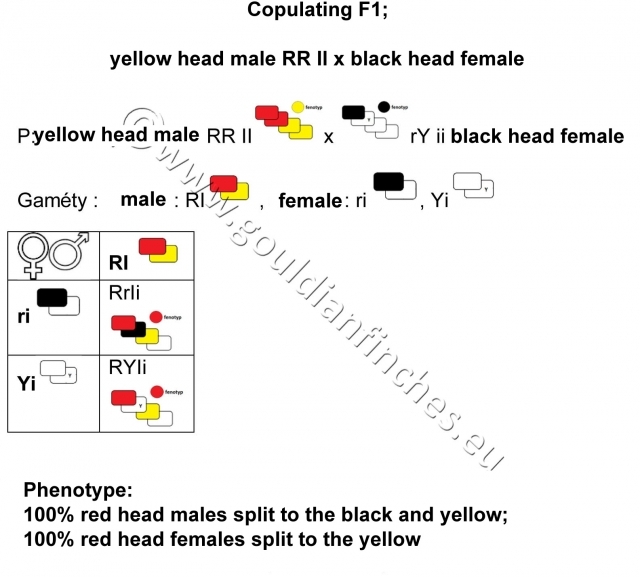
Breeding F2

Test breeding (breeding F1 generation with a recessive homozygote)
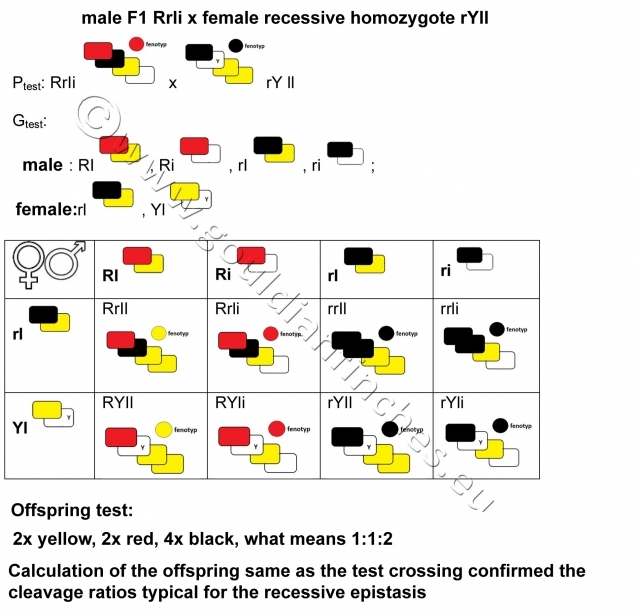
11.03.2014 -

this website www.gouldianfinchces.eu
Contact: