







Štruktúra peria a jeho sfarbenie
Perie a jeho sfarbenie

Perie
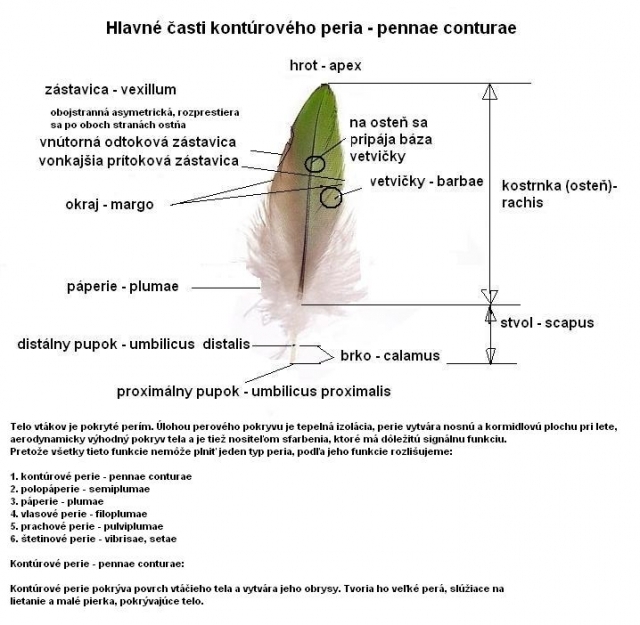
Telo vtákov je pokryté perím. Úlohou perového pokryvu je tepelná izolácia. Perie vytvára nosnú a kormidlovú plochu pri lete, aerodynamicky výhodný pokryv tela a je tiež nositeľom sfarbenia, ktoré má dôležitú signálnu funkciu. Perie sa na tele vtákov nepokrýva 100% ich pokožky, ale je rozdelené do „ostrovčekov“.


Všetky vyššie popísané funkcie nemôže plniť jeden typ peria, podľa jeho úlohy rozlišujeme:
1. kontúrové perie - pennaeconturae
2. polopáperie - semiplumae
3. páperie - plumae
4. vlasové perie - filoplumae
5. prachové perie - pulviplumae
6. štetinové perie – vibrisaesetae
1. Kontúrové perie - pennaeconturae
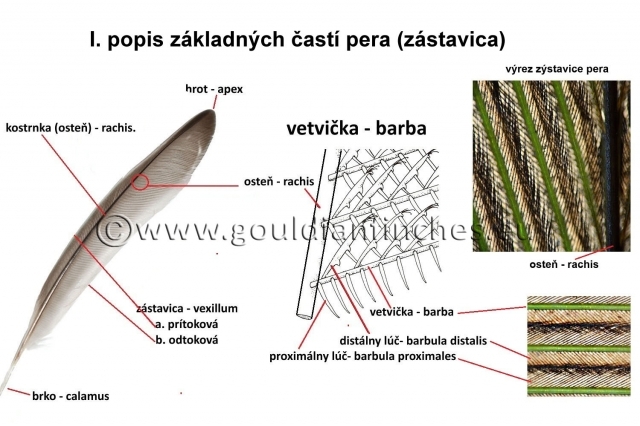
 Kontúrové perie pokrýva povrch vtáčieho tela a vytvára jeho obrysy. Tvoria ho veľké perá slúžiace na lietanie a malé pierka pokrývajúce telo. Os kontúrového peria tvorí stvol - scapus. Jeho bazálna časť brko - calamussedí pevne v pokožke. Brko je takmer duté, sú v ňom len zvyšky tzv. "duše". Malý otvor na konci brka sa nazýva proximálny pupok - umbilicusproximalis. V období vývinu pierka do neho týmto otvorom vstupuje zo škáry papila zásobovaná krvou. Tá časť stvola, po oboch stranách ktorej sa rozprestiera zástavica - vexillum, sa nazýva kostrnka - rachis. Kostrnka má na povrchu výživnú kôru - cortexa vnútro vyplnené vzduchom naplnenými komôrkami, ktoré sa nazývajú dreň - medulla. Na spodnej strane ostňa beží po celej jeho dĺžke ryha, ktorá v mieste prechodu ostňa do brka končí. Toto miesto sa nazýva distálny pupok – umbilicusdistalis. Časť zástavice na distálnom konci pera sa nazýva hrot - apex, ktorý na oboch stranách prechádza v okraj - margo. Mnohé kontúrové perá majú na hranici medzi kostrnkou a brkom umiestnené podpierko (bočné pierko) - hypopenna, ktoré má vlastnú podkostrnku - hyporachisa podzástavicu - hypovexillum. Podpierko je na ventrálnej strane pera a je spravidla omnoho menšie ako hlavné pierko. Chýba mnohým skupinám vtákov (Pelecaniformes, mnohým sovám - Strigiformes, spevavcom - Passeriformes). Naopak zreteľné je pri kurovitých – Galliformes a pri rode Emu a kazuároch je presne také dlhé ako kostrnka. U nich pravdepodobne slúži na zahustenie operenia. Zástavica letiek a bočných (okrajových) kormidlových pier je spravidla asymetrická. Rozlišujeme na nich užšiu a pevnejšiu vonkajšiu časť zástavice a jej širšiu mäkkšiu vnútornú časť.
Kontúrové perie pokrýva povrch vtáčieho tela a vytvára jeho obrysy. Tvoria ho veľké perá slúžiace na lietanie a malé pierka pokrývajúce telo. Os kontúrového peria tvorí stvol - scapus. Jeho bazálna časť brko - calamussedí pevne v pokožke. Brko je takmer duté, sú v ňom len zvyšky tzv. "duše". Malý otvor na konci brka sa nazýva proximálny pupok - umbilicusproximalis. V období vývinu pierka do neho týmto otvorom vstupuje zo škáry papila zásobovaná krvou. Tá časť stvola, po oboch stranách ktorej sa rozprestiera zástavica - vexillum, sa nazýva kostrnka - rachis. Kostrnka má na povrchu výživnú kôru - cortexa vnútro vyplnené vzduchom naplnenými komôrkami, ktoré sa nazývajú dreň - medulla. Na spodnej strane ostňa beží po celej jeho dĺžke ryha, ktorá v mieste prechodu ostňa do brka končí. Toto miesto sa nazýva distálny pupok – umbilicusdistalis. Časť zástavice na distálnom konci pera sa nazýva hrot - apex, ktorý na oboch stranách prechádza v okraj - margo. Mnohé kontúrové perá majú na hranici medzi kostrnkou a brkom umiestnené podpierko (bočné pierko) - hypopenna, ktoré má vlastnú podkostrnku - hyporachisa podzástavicu - hypovexillum. Podpierko je na ventrálnej strane pera a je spravidla omnoho menšie ako hlavné pierko. Chýba mnohým skupinám vtákov (Pelecaniformes, mnohým sovám - Strigiformes, spevavcom - Passeriformes). Naopak zreteľné je pri kurovitých – Galliformes a pri rode Emu a kazuároch je presne také dlhé ako kostrnka. U nich pravdepodobne slúži na zahustenie operenia. Zástavica letiek a bočných (okrajových) kormidlových pier je spravidla asymetrická. Rozlišujeme na nich užšiu a pevnejšiu vonkajšiu časť zástavice a jej širšiu mäkkšiu vnútornú časť.
Zástavica- vexillum
Zástavica musí byť hustá, pevná a zároveň poddajná, aby ju silný tlak vzduchu pri lietaní nerozbil. Po oboch stranách ostňa sú dva paralelné rady vetvičiek - barbae (rami) úzke, vedľa seba ležiace a vytvárajúce vlastnú plochu zástavice.
Vetvičky - barbae (rami)
Vetvičky sú na priereze silno stlačené, na užšej strane vetvičky je dorzálna hrana - cristadorsalisa oproti nej na spodnej strane ventrálna hrana – cristaventralis. Dorzálna a ventrálna hrana vetvičky môžu byť pri jednotlivých perách rozdielne silné. Napríklad pri mnohých letkách je silno rozšírená cristaventralis. Dreň vetvičky - medullanie je spojená s ostňom, ale začína sa na báze vetvičky. Vetvičky, nachádzajúce sa v ploche zástavice - parspennaceamajú po oboch stranách lúče - barbulae. Práve v keratíne a štruktúre vetvičiek (barba) sú prítomné pigmenty a tvorí sa tu tiež štrukturálna farba.
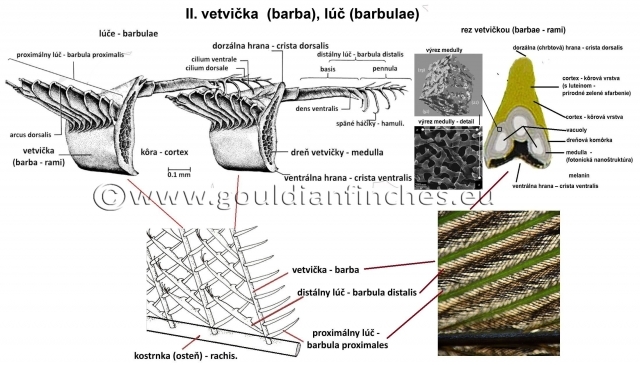
Lúče – barbulae
 Lúče sú sklonené smerom k hrotu vetvičky, takže s ňou zvierajú uhol menší ako 90°. Kompaktná bazálna časť distálneho lúča – barbuladistalis prechádza do apikálnej časti opatrenej spätnými háčikmi - hamuli. Preto sa tieto lúče nazývajú aj háčikovité. Proximálny lúč – barbulaproximalis nemá spätné háčiky a na konci vybieha do jemného hrotu. Na svojej dorzálnej strane sú proximálne lúče po celej svojej dĺžke zahnuté do oblúčika, preto sa nazývajú aj oblúčikovité. O tieto oblúčiky sú zachytené distálne lúče svojimi háčikmi. Šikmým postavením lúčov na vetvičke je dosiahnuté to, že háčiky každého distálneho (háčikového) lúča sú zachytené o niekoľko proximálnych (oblúčikovitých) lúčov. Pružnosť celého tohto systému sa zvyšuje aj tým, že distálne (háčikové) lúče sa môžu šmýkať po okrajoch proximálnych (oblúčikovitých) lúčov. Pri rôznych perách, ale aj v rôznych častiach toho istého pera môže byť spojenie jednotlivých lúčov prispôsobené rôznym požiadavkám, ktoré sú kladené na pero podľa jeho postavenia. Pri silnom tlaku vzduchu, ktorý pôsobí na letky pri lietaní, musí byť spevnená predovšetkým jeho koncová časť, aby nedochádzalo k veľmi rýchlemu rozbitiu pera. V lúčoch (barbulách) peria sa tiež nachádzajú pigmenty, a to najmä melaníny, ktoré môžu ovplyvniť celkové zafarbenie peria.
Lúče sú sklonené smerom k hrotu vetvičky, takže s ňou zvierajú uhol menší ako 90°. Kompaktná bazálna časť distálneho lúča – barbuladistalis prechádza do apikálnej časti opatrenej spätnými háčikmi - hamuli. Preto sa tieto lúče nazývajú aj háčikovité. Proximálny lúč – barbulaproximalis nemá spätné háčiky a na konci vybieha do jemného hrotu. Na svojej dorzálnej strane sú proximálne lúče po celej svojej dĺžke zahnuté do oblúčika, preto sa nazývajú aj oblúčikovité. O tieto oblúčiky sú zachytené distálne lúče svojimi háčikmi. Šikmým postavením lúčov na vetvičke je dosiahnuté to, že háčiky každého distálneho (háčikového) lúča sú zachytené o niekoľko proximálnych (oblúčikovitých) lúčov. Pružnosť celého tohto systému sa zvyšuje aj tým, že distálne (háčikové) lúče sa môžu šmýkať po okrajoch proximálnych (oblúčikovitých) lúčov. Pri rôznych perách, ale aj v rôznych častiach toho istého pera môže byť spojenie jednotlivých lúčov prispôsobené rôznym požiadavkám, ktoré sú kladené na pero podľa jeho postavenia. Pri silnom tlaku vzduchu, ktorý pôsobí na letky pri lietaní, musí byť spevnená predovšetkým jeho koncová časť, aby nedochádzalo k veľmi rýchlemu rozbitiu pera. V lúčoch (barbulách) peria sa tiež nachádzajú pigmenty, a to najmä melaníny, ktoré môžu ovplyvniť celkové zafarbenie peria.

Kontúrové perie – pennaeconturae môžeme podľa jeho použitia rozdeliť na:
letky – remiges

krovky – tectrices

kormidlové perá – rectrices

Perie ako optická štruktúra
Perie okrem svojej najdôležitejšej funkcie, teda zabezpečenie tepla a letu plní aj úlohu pri výbere partnera. Teda lepšie povedané je to jeho sfarbenie, ktoré upúta partnerku a môže tak prísť k „sobášu“ a odchovu mláďat. Odchov mláďat je najdôležitejšia funkcia každého živočíšneho druhu. Bez zabezpečenia reprodukcie by zanikol. Keďže je sfarbenie peria skutočne dôležité, povieme si o jeho tvorbe a vnímaní farieb viac. Je potrebné zdôrazniť, že perie je optická štruktúra, a tak svetlo a jeho vnímanie hrajú dôležitú úlohu. Pre pochopenie nasledujúcej kapitoly je potrebné si prečítať kapitolu „Svetlo, jeho vlastnosti a jeho vnímanie ľuďmi a vtákmi“.
Zloženie peria
Perie je tvorené z keratínu - vláknitého proteínu, ktorý sa tvorí v bunkách kože, ktoré formujú tvrdý, pružný a nerozpustný polymér. Na rozdiel od α - keratínu u cicavcov (pokožka a chlpy - vlasy), patrí keratín z peria, pazúrikov a zobáka vtákov k β - keratínom. Pierka, rohovité šupiny, pazúriky a zobák vtákov sú zložené z podtriedy β - keratínov, ktoré sú známe ako perové keratíny alebo φ – keratíny. Keratín peria obsahuje relatívne vysoké množstvo glycínovej, serínovej, prolínovej, leucinovej a glutamovej kyseliny. Keď porovnávame β - keratín peria s α - keratínom z chlpov cicavcov, pierka obsahujú relatívne menej methioninu a lyzínu (esenciálna aminokyselina) a vyšší obsah prolinu. Perie je teda tvorené β - keratínom, v ktorom sa môžu, alebo nemusia nachádzať pigmenty.
Farby peria
Perie je optická štruktúra a jeho sfarbenie je úzko spojené so svetlom. To, ako sa v perí svetlo láme, v štruktúre stavby peria a za prítomnosti pigmentov vytvára jeho sfarbenie, ktoré je viditeľné okom pozorovateľa.
Táto kapitola je rozdelená do 3 základných častí:
1. štruktúra peria,
2. pigmenty,
3. optická alebo štrukturálna farba.
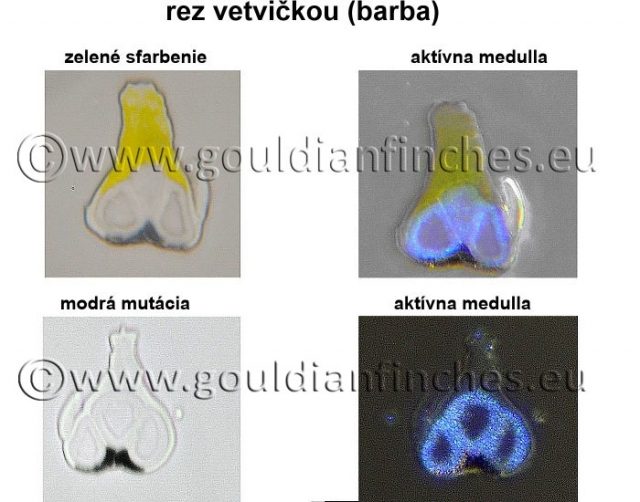
Sfarbenie peria amadín Gouldovej
Dva druhy melanínu :
- čierny a hnedý (eumelanín) - jeho zastúpenie vidíme na fotkách z peria čiernohlavých vtákov, kde je jasne v reze vidieť aj jeho intenzita uloženia v perí.
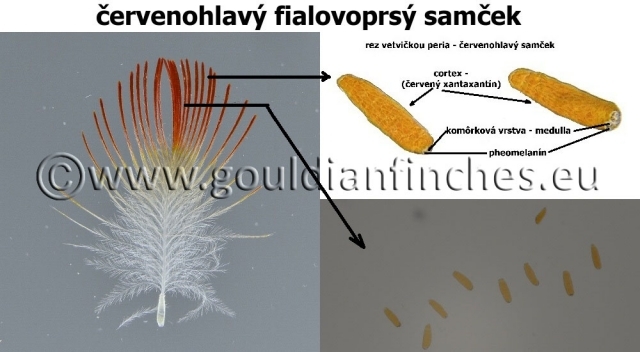
- hnedo - červený (pheomelanín). Na fotke z pŕs fialovoprsého vtáka je pheomelanín zodpovedný za vytvorenie štrukturálnej optickej farby a spolu s vlastnou farbou vytvárajú sfarbenie pŕs. V reze je vidieť jeho umiestnenie v perí.
Dva karotenoidy:
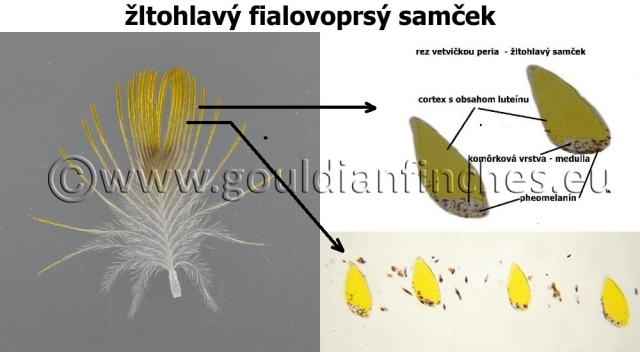
- žltý (luteín epoxid – alfa xanthofyl)*. Na fotke žltohlavého vtáka je ilustrované nielen rozloženie luteínu, ale v komôrkovej vrstve je prítomný tiež pheomelanín.
- červený (canthaxanthin (kantaxantin) – beta karotenoid)*. Jeho intenzita je vidieť na fotkách červenohlavých vtákov. Podobne ako u žltohlavých je prítomný aj pheomelanín, čo má za následok, že bieloprsé vtáky majú pre neprítomnosť pheomelanínu jasnejšie sfarbenie hlavy, čo je vidieť aj v štruktúre peria z makrofotografie.
*J. Wilson, R. Clark 1998
Podrobne o vzniku sfarbenia u vtákov píšem v kapitole „Ako vzniká sfarbenie“
Štrukturálne sfarbenie
Štrukturálne sfarbenie môže byť rozdelené na iridescentné a neiridescentné. Iridescentná farba sa mení s uhlom pohľadu či osvetlením, zatiaľ čo neiridescentné farby zostávajú rovnaké, nezávisle na uhle dopadu svetla.

Iridescentné farby (napr. menlivé farby s uhlom pohľadu alebo osvetlením;
Newton, 1704). Ide o interferenciu na tenkej vrstve (podobne ako u CD). Svetlo je odrážané od (drsného) povrchu peria, nie je teda lámané dovnútra. Niekoľko vrstiev keratínu, ktoré obsahujú melanínové zrná a pigmenty a tiež vzduchové (medzierky), môže byť naskladaných na seba tvoriac trojrozmernú mriežku vytvárajúcu viacvrstvové odrazové plochy.

Neiridescentné štrukturálne farby (modrá, fialová; nemeniace sa uhlom dopadu svetla ani uhlom pohľadu pozorovateľa) vtáčích pier sú tvorené súvislým rozptylom svetla v amorfnej (nepravidelne usporiadanej - kvázi usporiadanej) fotonickej nanoštruktúre β - keratínových medulárnych buniek a vzduchom vyplnených kanálikov vo vetvičke - barbe (rami) pierka s rôznym indexom lomu svetla. Súvislé rozptýlenie znamená, že fázové vzťahy medzi rozptýlenými svetelnými vlnami určujú výsledné viditeľné spektrum. Koherentný rozptyl obsahuje konštruktívnu interferenciu, difrakciu (ohyb svetla), lom na tenkej vrstve.
Fotonické nanoštruktúry
Známe nanoštruktúry vo vetvičke – barbe pera, patria do dvoch rozdielnych morfologických typov:

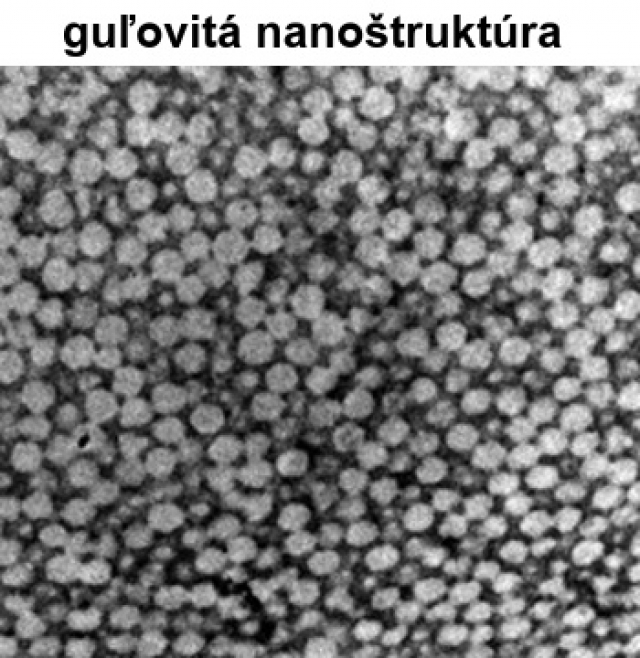 kanáliková nanoštruktúra – skladá sa z β - keratínových buniek tyčinkovitého tvaru a rozvetvených nepravidelných vzduchových kanálikov
kanáliková nanoštruktúra – skladá sa z β - keratínových buniek tyčinkovitého tvaru a rozvetvených nepravidelných vzduchových kanálikov
guľovitá nanoštruktúra – skladá sa z kruhových vzduchových dutiniek, ktoré sú ohraničené tenkými β - keratínovými tyčinkami, niekedy vzájomne popretkávanými.
Fotonické nanoštruktúry: môžu byť pravidelné štruktúry na veľkých pravidelných plochách alebo štruktúry neurčitého tvaru s odlišným indexom lomu v jednej, dvoch alebo troch osiach. Fotonické štruktúry môžu byť jedno, dvoj alebo trojrozmerné a teda sa líšia aj indexom lomu svetla. Fotonické nanoštruktúry sa líšia nielen v indexe lomu, čo je spôsobené ich architektúrou (1,2 alebo 3 rozmery), ale index lomu svetla vo fotonických štruktúrach sa líši tiež podľa ich morfológie.
Jednorozmerné (biofotonické) nanoštruktúry majú rôzne tenké filmy a viacvrstvové odrazové plochy (zrkadlá).
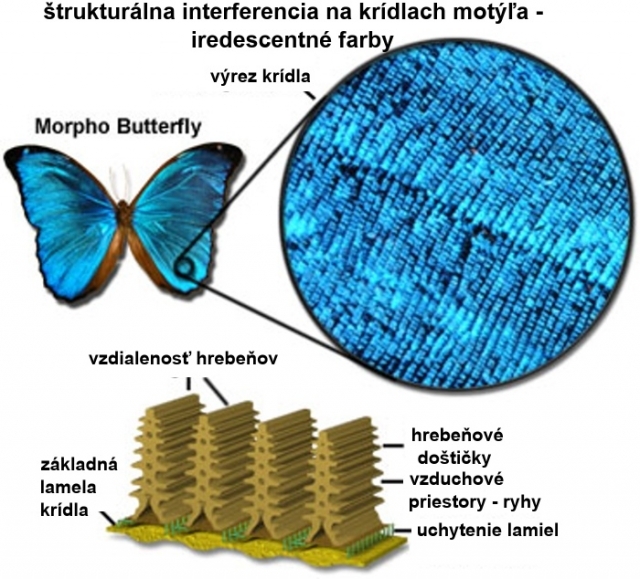
Dvojrozmerné štruktúry zahrňujú rady paralelných kolagénových vlákien, ktoré sa nachádzajú v koži vtákov, alebo napríklad hrebeňovité výrastky s doštičkami na krídlach motýľov.

Trojrozmerné štruktúry - patria sem aj špongiovité medulárne fotonické nanoštruktúry z barby pera vtákov. Trojrozmerné fotonické nanoštruktúry sú prepojené amorfné bikontinuálne matrice, ktoré sú primerane usporiadané v priestore vo všetkých troch rozmeroch (osiach) tak, že súvisle (koherentne) rozptyľujú svetlo.
Medulla v perí vtákov – fotonická nanoštruktúra
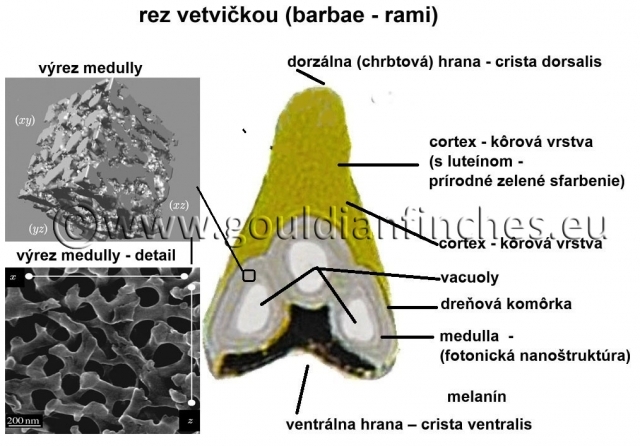 Zloženie a morfológia - skladá sa z buniek a kanálikov tvoriacich špongiovitú vrstvu (priemer keratínových vláken a vzduchových kanálikov), ktoré majú veľkosť približne 100 nm. Špongiovitá medulla, ktorú tvoria kanáliky naplnené vzduchom a bunky z keratínu, je umiestnená priamo pod cortexom, ktorý je z pevného keratínu a nad vrstvou melanínových zŕn, ktoré sú často ohraničené dutými vakuolami. Nanoštruktúra medully vytvárajúca štrukturálnu farbu je nepravidelná, bikontinuálna (t.j. všetky keratínové bunky a všetky kanáliky naplnené vzduchom sú medzi sebou prepojené, nejde teda o kryštalickú štruktúru) popretkávaná sieť keratínových buniek a vzduchových kanálikov. Táto fotonická nanoštruktúra je dostatočne periodická (striedajú sa bunky s kanálikmi) v príslušnom priestore vo všetkých troch osiach, aby sa v nej tvorila štrukturálna farba za pomoci interferencie z koherentných svetelných vĺn.
Zloženie a morfológia - skladá sa z buniek a kanálikov tvoriacich špongiovitú vrstvu (priemer keratínových vláken a vzduchových kanálikov), ktoré majú veľkosť približne 100 nm. Špongiovitá medulla, ktorú tvoria kanáliky naplnené vzduchom a bunky z keratínu, je umiestnená priamo pod cortexom, ktorý je z pevného keratínu a nad vrstvou melanínových zŕn, ktoré sú často ohraničené dutými vakuolami. Nanoštruktúra medully vytvárajúca štrukturálnu farbu je nepravidelná, bikontinuálna (t.j. všetky keratínové bunky a všetky kanáliky naplnené vzduchom sú medzi sebou prepojené, nejde teda o kryštalickú štruktúru) popretkávaná sieť keratínových buniek a vzduchových kanálikov. Táto fotonická nanoštruktúra je dostatočne periodická (striedajú sa bunky s kanálikmi) v príslušnom priestore vo všetkých troch osiach, aby sa v nej tvorila štrukturálna farba za pomoci interferencie z koherentných svetelných vĺn.
Utváranie a formovanie – špongiovité medulárne β - keratínové nanoštruktúry formujú svoje typické veľkosti a tvary fázovým delením počas proteínovej polymerizácie. V rastúcom pere sa nachádzajú živé keratínové medulárne bunky. V určitom stupni vývoja pera (v závislosti od druhu) začínajú medulárne bunky keratinizovať pozdĺž membránových ohraničení so susediacimi bunkami. Táto počiatočná periférna keratinizácia vytvára prepojené lineárne keratínové útvary, ktoré majú priemer desiatok nanometrov a viac (až do 100nm), a sú dlhé do 1 μm. Tieto procesy sa občas rozkonárujú alebo rozdvojujú vo veľmi malých uhloch (menej ako 30°) a vytvárajú rozvetvenú matricu. Keď sú medulárne bunky úplne dehydrované a mŕtve, vytvorené priestory v špongiovitej nanoštruktúre sa začínajú vypĺňať vzduchom vytvárajúc dielektrickú, kvázi - usporiadanú fotonickú nanoštruktúru, ktorá je zodpovedná za tvorbu transparentnej štrukturálnej farby v operení dospelých vtákov. Schnutie je základnou súčasťou keratinocytického zrenia. Všetky keratinocyty pera dosahujú svoje konečné funkcie až keď sú mŕtve a vysušené. Periférna keratinizácia v bunkových stenách, ktorá predchádza formovaniu dutín - kanálikov, má za následok znemožnenie zásobovania medulárnych buniek vodou, a tým nastáva úplné štrukturálne usporiadanie fotonickej nanoštruktúry do konečnej podoby.
Fotonické nanoštruktúry vytvárajúce štrukturálnu farbu sú veľmi podobné štruktúram, ktoré sa vytvorili samokompletovaním v čase fázového delenia nestabilnej zmesi, v procese známom ako spinodálna dekompozícia/rozklad (SD).
Tvorba štrukturálnej farby - neiredescentná (nemení sa uhlom dopadu svetla ani uhlom pozorovania)
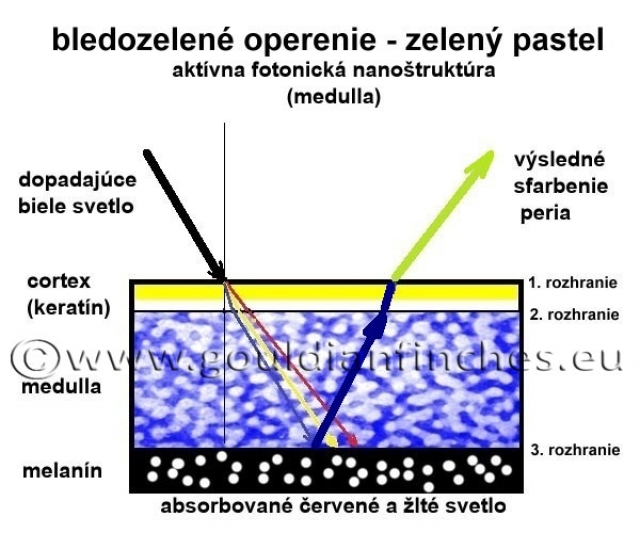
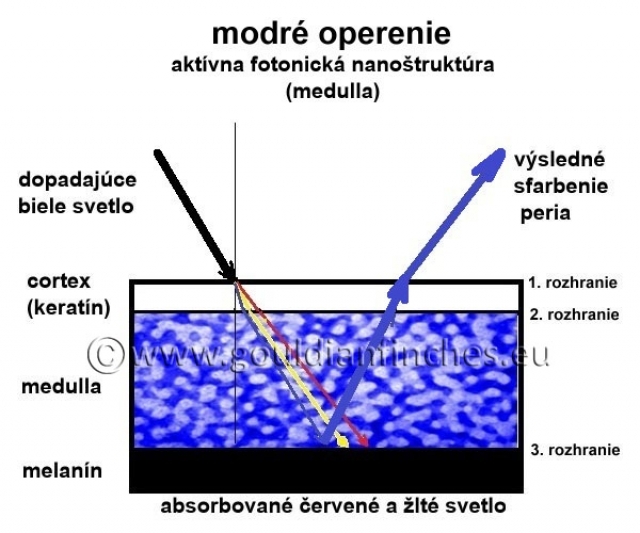
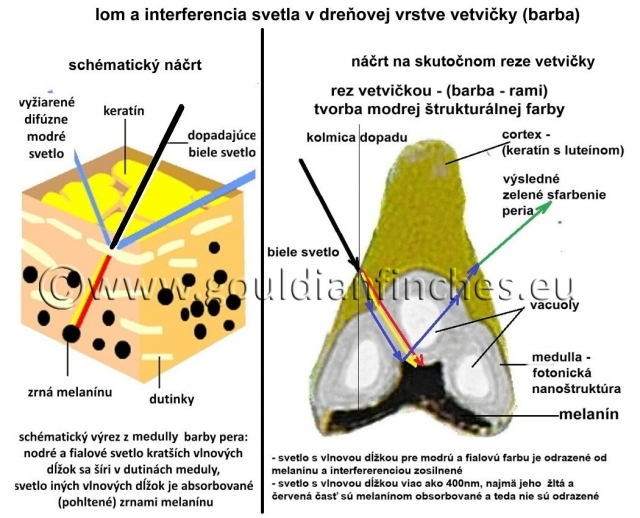 Odtieň vytvorenej štrukturálnej farby je závislý od rozmeru medulárnych buniek a rozmeru kanálikov (morfológia fotonickej nanoštruktúry). Celkové sfarbenie peria je ovplyvnené lúčmi (barbulae), ktoré obsahujú pigmenty a difúzne ako optické štruktúry rozptyľujú svetlo po celej ploche pera. Ak sú v komôrke dreňovej vrstvy prítomné zrná melanínu, absorbujú – pohlcujú svetlo iných vlnových dĺžok ako sú vlnové dĺžky pre modrú až fialovú časť spektra. Biele svetlo, zložené z celého spektra farieb, prechádza cez cortexbarby pera do medulárnej fotonickej nanoštruktúry. V zrnách melanínu je pohltená väčšina svetla vlnových dĺžok iných ako pre modrú až fialovú farbu. Svetlo s vlnovou dĺžkou modrej a fialovej farby je od melanínu odrazené, v medulle interferenciou zosilnené a vyžiarené cez cortex pera (zákon lomu a odrazu svetla).
Odtieň vytvorenej štrukturálnej farby je závislý od rozmeru medulárnych buniek a rozmeru kanálikov (morfológia fotonickej nanoštruktúry). Celkové sfarbenie peria je ovplyvnené lúčmi (barbulae), ktoré obsahujú pigmenty a difúzne ako optické štruktúry rozptyľujú svetlo po celej ploche pera. Ak sú v komôrke dreňovej vrstvy prítomné zrná melanínu, absorbujú – pohlcujú svetlo iných vlnových dĺžok ako sú vlnové dĺžky pre modrú až fialovú časť spektra. Biele svetlo, zložené z celého spektra farieb, prechádza cez cortexbarby pera do medulárnej fotonickej nanoštruktúry. V zrnách melanínu je pohltená väčšina svetla vlnových dĺžok iných ako pre modrú až fialovú farbu. Svetlo s vlnovou dĺžkou modrej a fialovej farby je od melanínu odrazené, v medulle interferenciou zosilnené a vyžiarené cez cortex pera (zákon lomu a odrazu svetla).
Mechanizmus vzniku rôzneho sfarbenia peria
Zelená farba: Ak vyžiarené modré štrukturálne svetlo prechádza cez cortex pera, v ktorom sa nachádza pigment - žltý luteín, výslednou farbou pera bude zelená (modrá + žltá). Zelená farba je teda súčtom pigmentového sfarbenia, a to žltého luteínu spolu s modrou štrukturálnou farbou. Intenzita a odtieň zelenej sa môžu líšiť od množstva pigmentu v cortexe pera, ako aj od odtieňa modrej štrukturálnej farby.


Modrá farba: Ak sa v cortexe pigment nenachádza, výsledná farba bude modrá tak, ako bola vytvorená v medulle interferenciou. Modré sfarbenie teda nie je sfarbením, kde by bolo niečo pridané, ale práve naopak, v perí chýba žltý luteín, či iné farbivo, ktoré by ovplyvňovalo celkové sfarbenie peria. Keďže je cortex „prakticky“ bezfarebný, z komôrkovej vrstvy je vyžiarené modré svetlo, ktoré určuje sfarbenie peria. Zvyčajne je však pero modro sfarbené z vrchnej časti a v spodnej má farbu prítomného pigmentu.

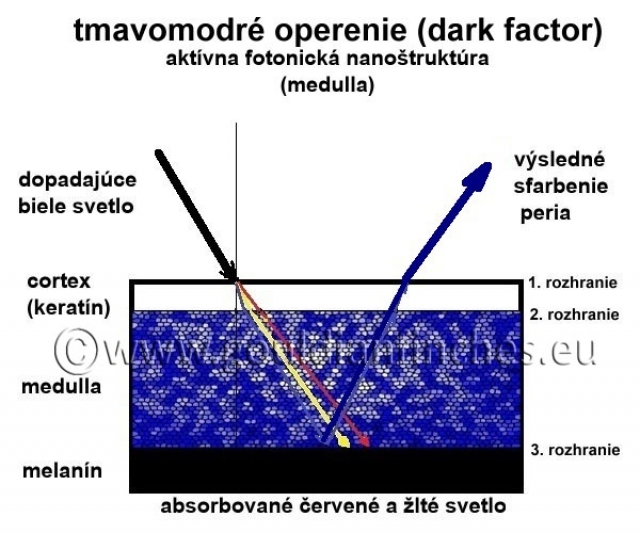
Dark faktor: Zmena hustoty a veľkosti medulárnych buniek či zmena veľkosti kanálikov spôsobuje zmenu indexu lomu svetla, čo má za následok, že výsledné modré štrukturálne sfarbenie bude mať iný odtieň ako pri nezmenených pomeroch v medulárnej fotonickej nanoštruktúre (medulle). Zmena štruktúry v medulle spôsobuje napríklad efekt tmavnutia peria, tzv. dark - faktor (prichádza k zhusteniu, zúženiu kanálikov). Tmavnutie v tomto prípade nespôsobuje zmena množstva tmavého pigmentu, ale vytvorenie modrej farby tmavšieho odtieňa.
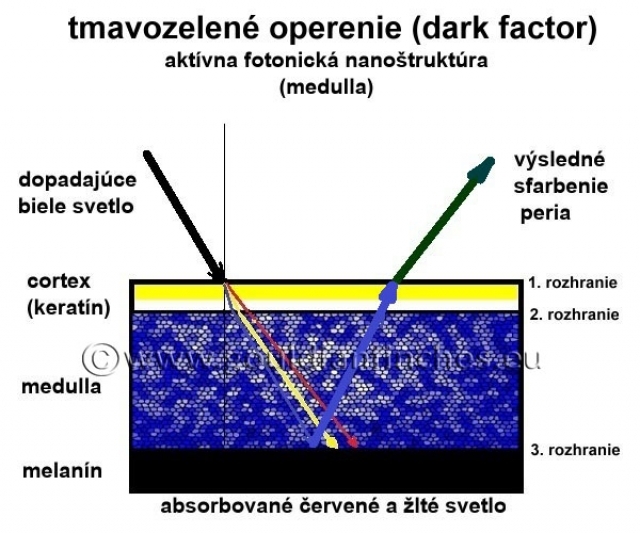
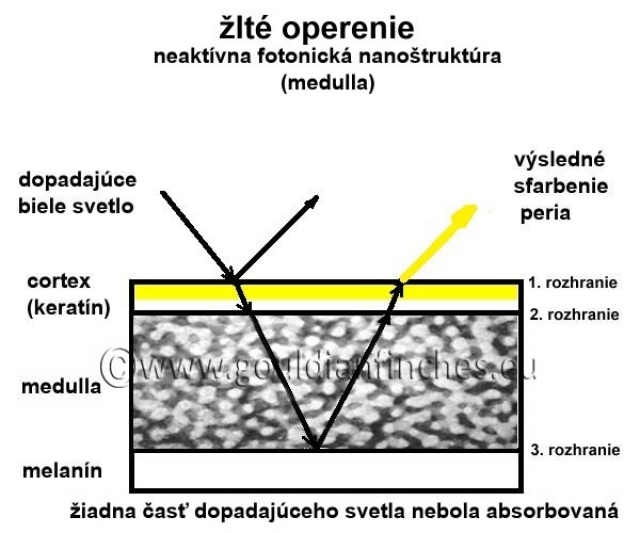
Perie bez štrukturálnej farby: Ak v medulle nie je prítomný melanín, žiadna časť bieleho svetla nemôže byť ním absorbovaná. V tomto prípade zvyčajne dochádza tiež k zmene veľkosti medulárnych buniek (rozmer viac ako 400 nm), za týchto podmienok sa nevytvorí ani modré štrukturálne svetlo (rozmer buniek viac ako
400 nm je väčší ako vlnová dĺžka pre modré svetlo) a cez cortex pera je teda spätne vyžiarené biele svetlo. V tomto prípade nastávajú 2 možnosti:
1. biela farba: Ak nie je cortex sfarbený, tak výslednou farbou peria bude biela,


iná farba: Ak je v cortexe prítomný nejaký pigment (melanín, carotenoid, ich kombinácia alebo psitacín), tak výsledná farba peria bude mať farbu melanínu nachádzajúceho sa v cortexe



Špecifikum je sfarbenie žltej a červenej hlavy u amadín Gouldovej, komôrková časť je tak malá, že výsledné sfarbenie je ovplyvnené prakticky len pigmentom obsiahnutým v cortexe pera. Aj keď je však komôrková vrstva malá, nachádza sa v nej určité množstvo melanínu, ktorý spôsobuje namiesto žltého sfarbenia hlavy sfarbenie oranžové. Pri bieloprsých vtákoch, kde nastáva úbytok melanínu – pheomelanínu je výsledné sfarbenie peria jasnejšie, menej oranžové a červené je žiarivejšie.
Odoberanie vajíčok a ich následná inkubácia
02.11.2016 - Kniha o chove amadín Gouldovej.
http://gfinches2.com/
http://www.gouldamadine.de/
http://www.novaexota.eu/
http://www.epapousek.cz/

stránok www.gouldianfinchces.eu
Kontakt: